 图片附件: 66943310.jpg (2013-6-27 11:24, 54.24 KB) / 该附件被下载次数 9
图片附件: 66943310.jpg (2013-6-27 11:24, 54.24 KB) / 该附件被下载次数 9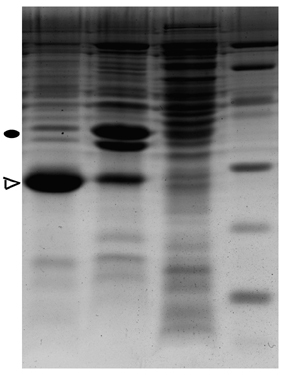
 。我用的冰浴,超声10S,间隔15S,超声时长30min。发现有很多泡沫。超声后我进行全菌蛋白SDS-PAGE电泳,发现上清和沉淀以及全菌没有表达,原因是什么?哪位大神能帮忙指导一下?感激
。我用的冰浴,超声10S,间隔15S,超声时长30min。发现有很多泡沫。超声后我进行全菌蛋白SDS-PAGE电泳,发现上清和沉淀以及全菌没有表达,原因是什么?哪位大神能帮忙指导一下?感激 ,可以1邮箱联系415977124@qq.com图片附件: [上清全菌检测] 微信图片_20200611115100.png (2020-6-11 11:54, 486.15 KB) / 该附件被下载次数 2
,可以1邮箱联系415977124@qq.com图片附件: [上清全菌检测] 微信图片_20200611115100.png (2020-6-11 11:54, 486.15 KB) / 该附件被下载次数 2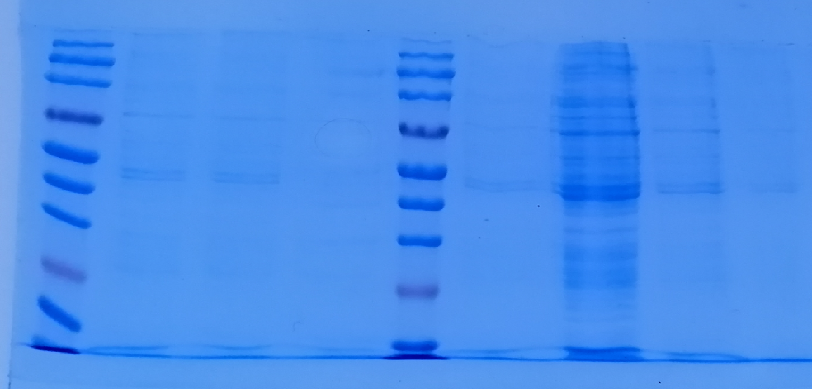
| 欢迎光临 分析测试百科 (http://bbs.antpedia.com/) | Powered by Discuz! 5.5.0 |