 图片附件: 43085935.jpg (2013-7-19 16:02, 59.85 KB) / 该附件被下载次数 10
图片附件: 43085935.jpg (2013-7-19 16:02, 59.85 KB) / 该附件被下载次数 10 图片附件: 10980078.jpg (2013-7-19 16:03, 20.96 KB) / 该附件被下载次数 14
图片附件: 10980078.jpg (2013-7-19 16:03, 20.96 KB) / 该附件被下载次数 14 图片附件: 38239610.jpg (2013-7-19 16:03, 18.14 KB) / 该附件被下载次数 16
图片附件: 38239610.jpg (2013-7-19 16:03, 18.14 KB) / 该附件被下载次数 16 图片附件: 12684741.gif (2013-7-19 16:03, 22.84 KB) / 该附件被下载次数 14
图片附件: 12684741.gif (2013-7-19 16:03, 22.84 KB) / 该附件被下载次数 14 图片附件: 41388716.snap.jpg (2013-7-19 16:10, 40.42 KB) / 该附件被下载次数 9
图片附件: 41388716.snap.jpg (2013-7-19 16:10, 40.42 KB) / 该附件被下载次数 9 图片附件: 79862920.jpg (2013-7-19 16:34, 18.39 KB) / 该附件被下载次数 7
图片附件: 79862920.jpg (2013-7-19 16:34, 18.39 KB) / 该附件被下载次数 7 图片附件: 55655793.gif (2013-7-19 16:34, 4.97 KB) / 该附件被下载次数 17
图片附件: 55655793.gif (2013-7-19 16:34, 4.97 KB) / 该附件被下载次数 17 图片附件: 70526190.jpg (2013-7-19 16:35, 22.19 KB) / 该附件被下载次数 8
图片附件: 70526190.jpg (2013-7-19 16:35, 22.19 KB) / 该附件被下载次数 8 图片附件: 47954246.jpg (2013-7-19 16:41, 26.09 KB) / 该附件被下载次数 23
图片附件: 47954246.jpg (2013-7-19 16:41, 26.09 KB) / 该附件被下载次数 23 图片附件: 43302075.snap.jpg (2013-7-19 16:45, 17.97 KB) / 该附件被下载次数 22
图片附件: 43302075.snap.jpg (2013-7-19 16:45, 17.97 KB) / 该附件被下载次数 22 图片附件: 74496872.snap.jpg (2013-7-19 17:01, 101.18 KB) / 该附件被下载次数 13
图片附件: 74496872.snap.jpg (2013-7-19 17:01, 101.18 KB) / 该附件被下载次数 13 图片附件: 14356795.snap.jpg (2013-7-19 17:02, 73.5 KB) / 该附件被下载次数 16
图片附件: 14356795.snap.jpg (2013-7-19 17:02, 73.5 KB) / 该附件被下载次数 16 图片附件: 84227336.jpg (2013-7-19 17:05, 39.92 KB) / 该附件被下载次数 33
图片附件: 84227336.jpg (2013-7-19 17:05, 39.92 KB) / 该附件被下载次数 33 图片附件: 94141251.jpg (2013-7-19 17:06, 13.84 KB) / 该附件被下载次数 13
图片附件: 94141251.jpg (2013-7-19 17:06, 13.84 KB) / 该附件被下载次数 13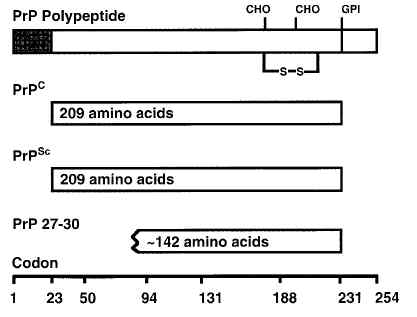 图片附件: 59059223.jpg (2013-7-19 17:07, 6.1 KB) / 该附件被下载次数 13
图片附件: 59059223.jpg (2013-7-19 17:07, 6.1 KB) / 该附件被下载次数 13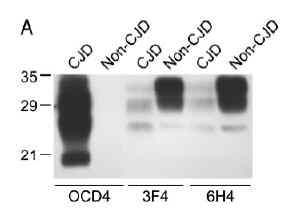 图片附件: 85851396.jpg (2013-7-19 17:08, 10.67 KB) / 该附件被下载次数 12
图片附件: 85851396.jpg (2013-7-19 17:08, 10.67 KB) / 该附件被下载次数 12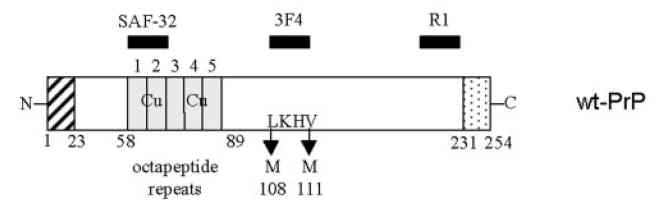 图片附件: 16564676.jpg (2013-7-19 17:09, 40.04 KB) / 该附件被下载次数 12
图片附件: 16564676.jpg (2013-7-19 17:09, 40.04 KB) / 该附件被下载次数 12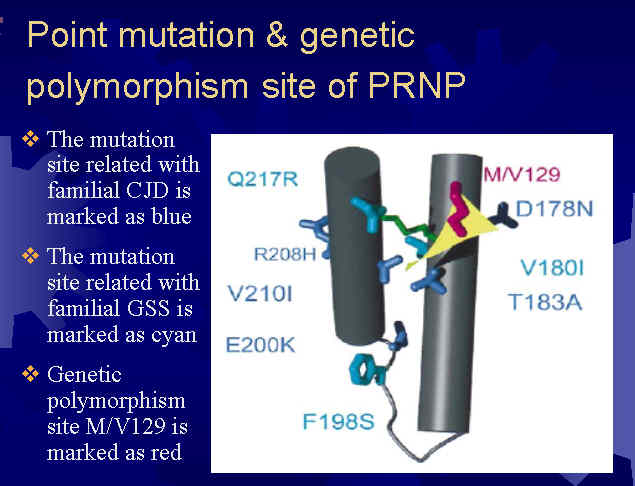 图片附件: 57064682.jpg (2013-7-19 17:10, 26.37 KB) / 该附件被下载次数 17
图片附件: 57064682.jpg (2013-7-19 17:10, 26.37 KB) / 该附件被下载次数 17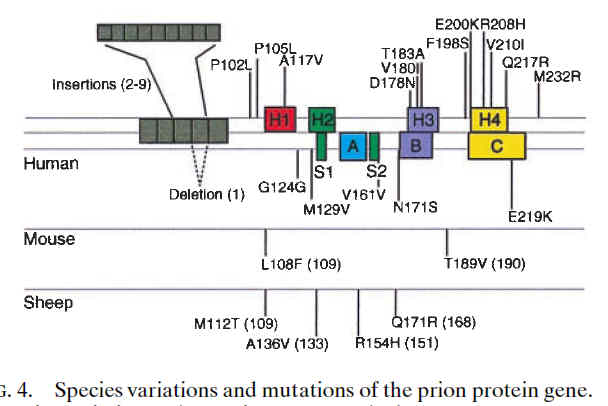 图片附件: 30395103.jpg (2013-7-19 17:11, 28.49 KB) / 该附件被下载次数 16
图片附件: 30395103.jpg (2013-7-19 17:11, 28.49 KB) / 该附件被下载次数 16 图片附件: 72507910.jpg (2013-7-19 17:11, 32.56 KB) / 该附件被下载次数 16
图片附件: 72507910.jpg (2013-7-19 17:11, 32.56 KB) / 该附件被下载次数 16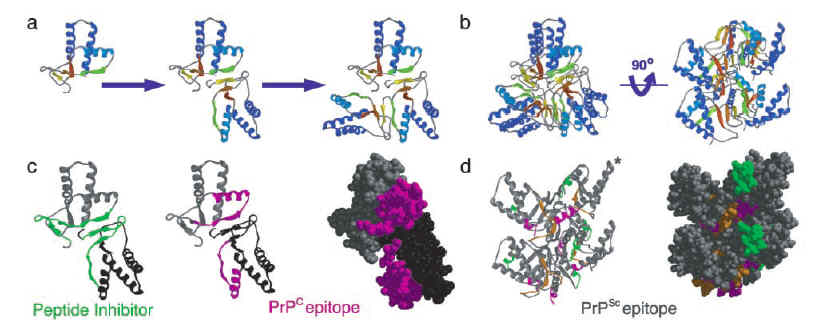 图片附件: 73688700.snap.jpg (2013-7-19 17:13, 93.72 KB) / 该附件被下载次数 22
图片附件: 73688700.snap.jpg (2013-7-19 17:13, 93.72 KB) / 该附件被下载次数 22
| 欢迎光临 分析测试百科 (http://bbs.antpedia.com/) | Powered by Discuz! 5.5.0 |