 图片附件: 80610355.jpg (2016-4-14 11:11, 46.66 KB) / 该附件被下载次数 167
图片附件: 80610355.jpg (2016-4-14 11:11, 46.66 KB) / 该附件被下载次数 167 图片附件: 53866002.snap.jpg (2016-4-14 11:12, 98.62 KB) / 该附件被下载次数 135
图片附件: 53866002.snap.jpg (2016-4-14 11:12, 98.62 KB) / 该附件被下载次数 135 图片附件: 35546607.snap.jpg (2016-4-14 11:14, 53.53 KB) / 该附件被下载次数 118
图片附件: 35546607.snap.jpg (2016-4-14 11:14, 53.53 KB) / 该附件被下载次数 118 图片附件: 73817993.snap.jpg (2016-4-14 11:31, 94.41 KB) / 该附件被下载次数 128
图片附件: 73817993.snap.jpg (2016-4-14 11:31, 94.41 KB) / 该附件被下载次数 128 图片附件: 81970776.snap.jpg (2016-4-14 14:21, 97.9 KB) / 该附件被下载次数 10
图片附件: 81970776.snap.jpg (2016-4-14 14:21, 97.9 KB) / 该附件被下载次数 10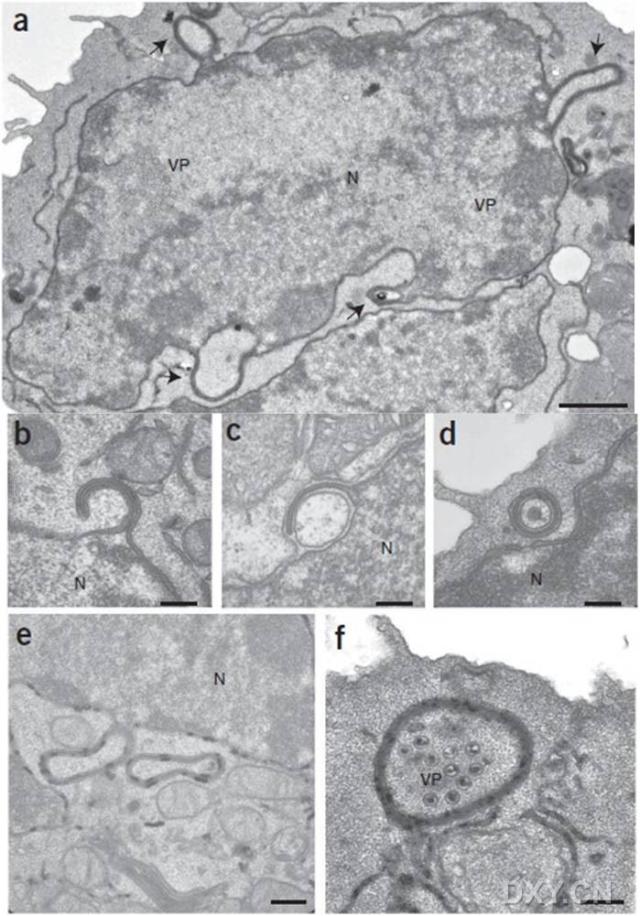 图片附件: 27402224.jpg (2016-4-14 14:25, 56.72 KB) / 该附件被下载次数 12
图片附件: 27402224.jpg (2016-4-14 14:25, 56.72 KB) / 该附件被下载次数 12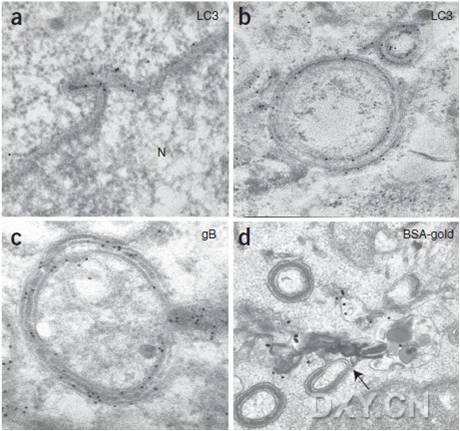 图片附件: 43170451.jpg (2016-4-14 15:39, 45.11 KB) / 该附件被下载次数 13
图片附件: 43170451.jpg (2016-4-14 15:39, 45.11 KB) / 该附件被下载次数 13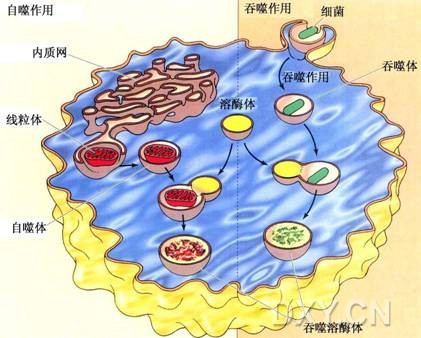 图片附件: 60289054.snap.jpg (2016-4-14 16:48, 155.45 KB) / 该附件被下载次数 8
图片附件: 60289054.snap.jpg (2016-4-14 16:48, 155.45 KB) / 该附件被下载次数 8 图片附件: 28570241.snap.jpg (2016-4-14 16:56, 77.56 KB) / 该附件被下载次数 3
图片附件: 28570241.snap.jpg (2016-4-14 16:56, 77.56 KB) / 该附件被下载次数 3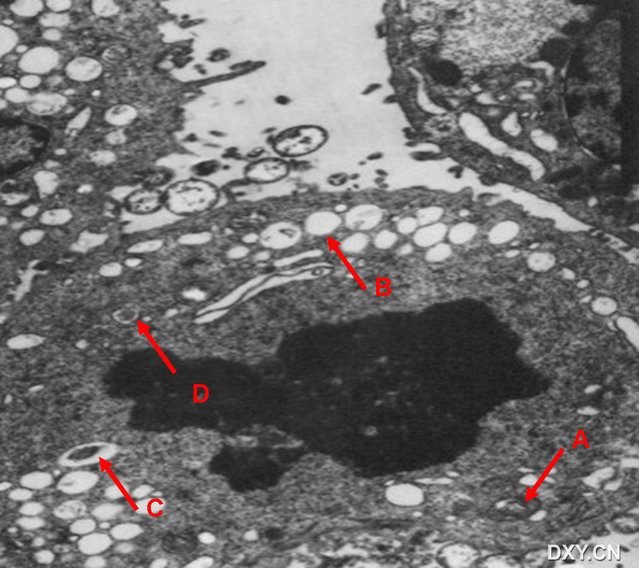 图片附件: 16068692.jpg (2016-4-15 11:04, 68.5 KB) / 该附件被下载次数 9
图片附件: 16068692.jpg (2016-4-15 11:04, 68.5 KB) / 该附件被下载次数 9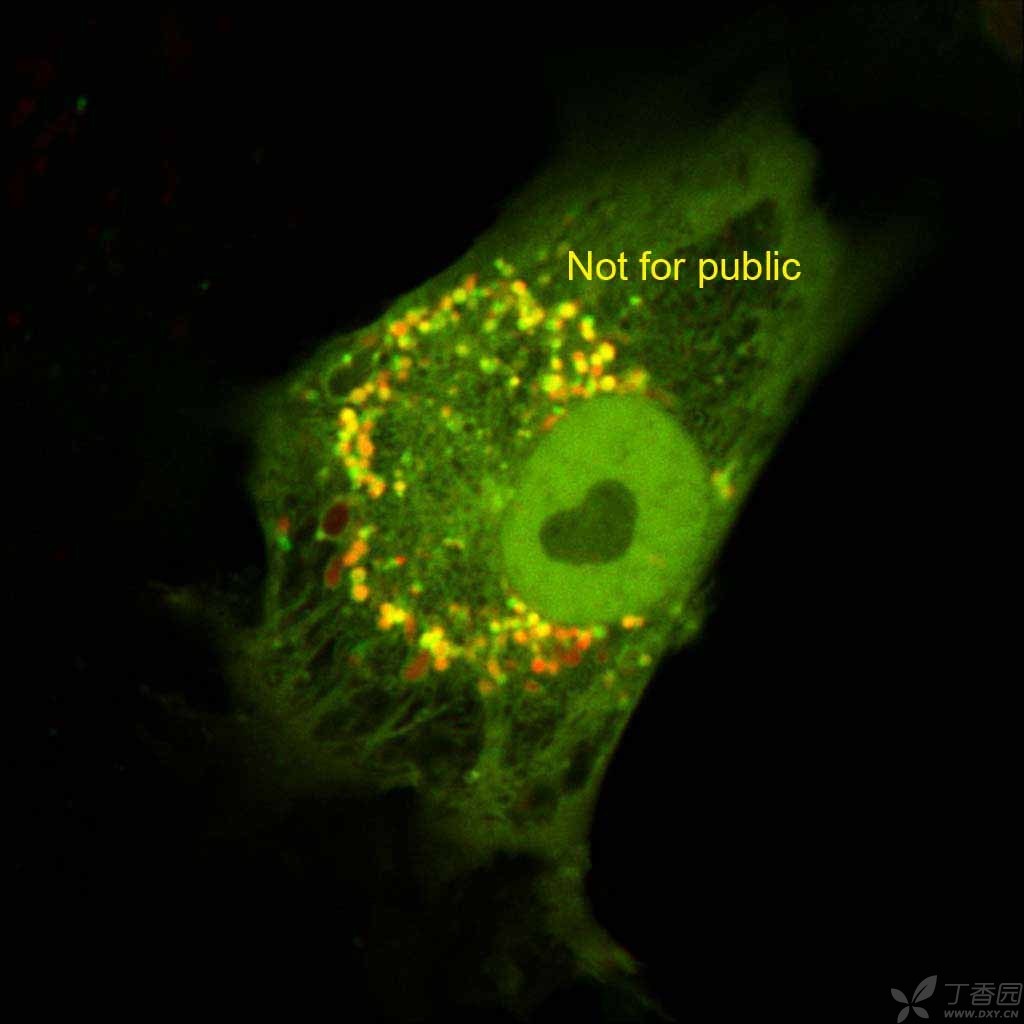 图片附件: 47881310.jpg (2016-4-15 11:07, 5.52 KB) / 该附件被下载次数 13
图片附件: 47881310.jpg (2016-4-15 11:07, 5.52 KB) / 该附件被下载次数 13 图片附件: 79041042.png (2016-4-15 11:12, 22.75 KB) / 该附件被下载次数 15
图片附件: 79041042.png (2016-4-15 11:12, 22.75 KB) / 该附件被下载次数 15 图片附件: 68507618.png (2016-4-15 11:17, 56.47 KB) / 该附件被下载次数 12
图片附件: 68507618.png (2016-4-15 11:17, 56.47 KB) / 该附件被下载次数 12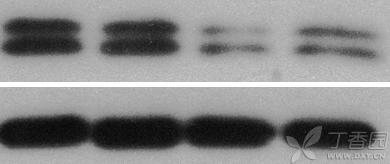
| 欢迎光临 分析测试百科 (http://bbs.antpedia.com/) | Powered by Discuz! 5.5.0 |